Karen Launchbaugh, Charles A. Taylor, Erika Straka and Robert Pritz
Introduction
In livestock management on rangelands we are often reminded that forage availability ” runs the show”. It seems we are always short of “forage”. Forage shortages can be solved by trucking in forage from somewhere else or converting the juniper hills into productive grasslands and shrublands by machine, fire, or chemical. These are expensive alternatives! Another solution might lie in our own back yards. Perhaps all that juniper could be converted into forage. To convert juniper from a weed to a feed must first answer the question: “Do animals get any nutrients out of juniper when they eat it?”.
Few livestock producers would worry about juniper invasion if juniper was a nutritious and palatable forage. Conversely, juniper in central Texas would be a less successful invader if it were highly palatable to herbivores. Therefore our second question must be: “Why is juniper unpalatable to most herbivores?”. The answer to these questions lies in understanding the biochemical composition of juniper and how herbivores process these biochemicals once they are ingested.
Chemical Composition of Juniper
How does juniper stack up against other range forage, from a nutritional standpoint? This depends on what season you are interested in. Juniper has a moderate amount of crude protein, phosphorus, and digestible organic matter (Table 1; Based on Huston et al. 1981). During the growing season, green grasses and forbs generally contain more energy and nutrients than juniper. However, as the season progresses into fall the herbaceous plants (grasses and forbs) loose much of their nutritional value while the nutrient and energy content of juniper remains relatively constant. Therefore, juniper could be a relatively good forage in fall and winter based on its nutrient content.
| Table 1. Chemical Composition of
Redberry Juniper Foliage |
|
| Nutrient | Percent of Dry Matter |
| Water | 46 to 56 |
| Ash | 4 to 6 |
| Cell Wall | 34 to 37 |
| Phosphorus | .09 to .17 |
| Crude Protein | 6 to 9 |
| Digestible Organic Matter | 57 to 66 |
| Components other than water are expressed on a dry matter basis.
(Data from Huston et al.. 1981) |
|
If juniper is relatively nutritious, then why is it considered marginal forage for livestock and wildlife? A closer look reveals that juniper contains chemical compounds that are detrimental to herbivores and decrease the forage value of juniper. Many range plants contain compounds that are toxic or detrimental to herbivores; these compounds are called allelochemicals. The allelochemicals in juniper are a specific group of compounds called volatile or essential oils. The properties of essential oils are described in the chapter by Taylor and associates of this proceedings.
Why is Juniper Unpalatable to Most Herbivores?
Herbivores may dislike juniper simply because it is not as nutritious as other forage. Herbivores can quite effectively distinguish between plants that differ in digestible energy and nutrients (Provenza 1995). However, as discussed earlier, juniper is moderately nutritious. Therefore, we assume that the reason herbivore dislike juniper is because of the essential oils.
Early studies on juniper consumption at the Sonora Experiment Station showed that when the percent of juniper in a mixed diet increased, the consumption of the diet by goats and sheep decreased. Curiously, subsequent research showed that when juniper is dried and ground, releasing most of the essential oils, it becomes almost entirely palatable (Fig. 1; Taylor et al. 1994). When blueberry (Juniperus ashei) and redberry juniper (Juniperus pinchotii) were added to an alfalfa meal, cottonseed meal and sorghum grain diet at rates of 0, 6.25, 12.5, and 25% it had no effect on the consumption of the diet, except at the highest level of added redberry juniper.
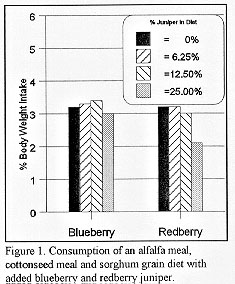{kind=link}
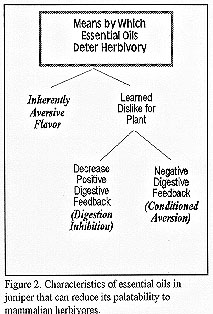
It is not known exactly how essential oils reduce the palatability of juniper. We can, however, hypothesize a limited number of ways in which the biochemical composition of any plant influences its probability of being grazed (Fig. 2).
{kind=link}
Inherently Aversive Flavor
Why is juniper unpalatable to livestock? Many people would say, “because it tastes bad”. It is possible that the strong flavor of juniper is inherently offensive to mammalian herbivores. Many scientists consider the ill flavor of some plants to be an important defense against herbivory (Laycock et al. 1988, Lindroth 1989). A significant number of plant allelochemicals are bitter or otherwise unpleasant to taste; at least to humans. Most herbivores initially avoid plants with strong flavors (Launchbaugh and Provenza 1994) or bitter flavors (Garcia and Hankins 1975). It may improve the survival of herbivores if they instinctively avoid strong or bitter flavors in plants because many plant toxins possess a bitter flavor (Garcia and Hankins 1975). However, the innate avoidance of bitter flavors is not universal and strong preferences can be formed to bitter-tasting foods when ingestion is followed by positive digestive consequences. Many foods eaten by humans, in fact, require a certain bitterness to confer palatability (e.g., coffee, beer, chocolate, cheddar cheese; Molyneux and Ralphs 1992). Additionally, livestock are often observed eating plants that are intensely bitter smelling or tasting to humans. The flavor of juniper may decrease its palatability, but flavor alone is probably not sufficiently deterrent to herbivores unless the tasting is followed by negative ingestive consequences.
Conditioned Aversions Based on Negative Digestive Feedback
If animals are not born with a dislike for the flavor of juniper then they learn to dislike juniper through the formation of a conditioned flavor aversion (CFA). When an herbivore encounters a new plant, it generally eats only a small amount of the plant. If illness does not follow ingestion, animals generally increase intake of the new plant over time. However, if the herbivore becomes ill after eating a new plant it forms a dislike for the plant, called a conditioned aversion (Fig. 3).
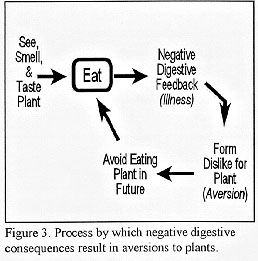{kind=link}
According to this theory, herbivores dislike juniper because it makes them ill when they eat it. To test this effect, we first distilled the essential oils out of redberry juniper leaves. We then fed 24 goats 300 grams (about 4 cups) of oats. After the goats had eaten the oats we gave them a bolus containing a low, medium, or high amount of essential oils. Two days later, we offered the goats oats again and measured how much they ate. We found that goats receiving the high amount of essential oils did not eat the oats; apparently because they became ill from the essential oils and formed an aversion to oats.
Different plant have different mixtures of specific terpenes in their essential oil fraction. Which terpenes are present and in what proportions probably influences the degree of illness that animal experiences after consumption. An example is that blueberry juniper is eaten more readily by goats than redberry juniper, probably because of the characteristic profiles of essential oils (Straka 1993, Riddle et al. 1996)
Digestion Inhibition
Another reason why livestock may find juniper unpalatable is that essential oils have been shown to inhibit the growth of rumen and caecal/colon microbes (Nagy et al. 1964, Oh et al. 1967). The antimicrobial and antifungal properties of essentials oils have been recognized for hundreds of years and are the basis of pine-scented anti-bacterial household cleaners of today. This antimicrobial effect of essential oils can be quite detrimental to ruminants (such as cows, sheep, goats and deer) and hind-gut fermentors (such as horses and rabbits) because these animals rely on microbes in their gut to breakdown and digest the fibrous portions of range forage. Reduced activity of microbes in the digestive tract would lead to decreased diet digestibility. Plants of low digestibility are usually less palatable than those of higher digestibility (Provenza 1995). Deer even prefer feed containing essential oils with low antimicrobial properties over those with strong antimicrobial properties (Schwartz et al. 1980).
The antimicrobial effect of essential oils has been demonstrated for several essential oil-containing plants and for several herbivore species including cows, goats, sheep, and deer. However, more recent work indicates that the monoterpene concentration in the rumens of deer eating sagebrush is likely too low to interfere greatly with microbial digestion (Cluff et al. 1982). It is not yet clear why ingested levels of monoterpenes are not always reflected in high rumen terpene concentrations. However, monoterpenes from ingested foliage may be rapidly absorbed across the rumen wall or volatilized before reaching the rumen through mastication or belching (Cluff et al. 1982, Gershenzon and Croteau 1991).
Why Do Some Animals Like Juniper More Than Others?
When given fresh juniper in pens, goats eat more than sheep which eat more than cows per unit of body weight (Straka 1993). And, among goats, Angoras generally eat more than Spanish goats (Ridell et al. 1996, Pritz et al. 1997). Unfortunately, we are not sure why this occurs. One explanation is that goats have a greater ability to detoxify the juniper essential oils than cattle or sheep. This may be evidenced by the fact that goats have bigger livers (per unit of body weight) than sheep or cows (Hofmann 1988). Furthermore, when we examined liver activity in Spanish and Angora goats eating juniper we observed less liver damage in the Spanish goats than Angoras (Pritz et al. 1997). This suggests that Spanish goats can eat more juniper than Angoras because they are more able to detoxify and excrete the essential oils.
There may be other explanations for differences between breeds and species. It is possible that goats and deer have different rumen microbes than cattle and sheep and that these microbes are more effective at detoxifying essential oils. It is also possible that deer and goats chew juniper more thoroughly than cattle and sheep releasing more of the essential oils before the juniper is swallowed. Finally, juniper may have equally negative effects on all breeds and species but animals may differ in their ability to tolerate the negative effects. In other words, goats and deer may simply be more willing or able to suffer illness from eating juniper than cattle and sheep.
How Do Animals Cope With Plant Allelochemcials?
Once juniper is ingested, the essential oils are liberated from the forage and they can either be detoxified by rumen microbes or enter the animal’s body. There are many plant toxins that are detoxified by rumen microbes and made benign to the ruminant animals. For example, rumen microbes in sheep detoxify oxalates from halogeton by forming insoluble calcium oxalates, which are excreted in the feces (Allison 1978). It is very possible that some species of rumen microbes have evolved the ability to detoxify the essential oils of juniper. However, scientists have not yet identified microbes that effectively modify essential oils.
Monoterpenes that are not detoxified by rumen microbes are absorbed across the rumen wall or through the small intestine. The monoterpenes are then transported to the liver and other tissues where they enter a series of reaction mediated by multi-function oxidases (MFO’s). These oxidase systems are important because they convert the terpenes into compounds that are water soluble (Brattsten 1979). This is an important reaction because mammals cannot easily rid toxins from their system unless they can be dissolved in water and pass out of the animal through urine or feces.
If the ingested terpenes are not removed from the system by detoxification in the rumen, liver, or other tissue they can have several detrimental effects on the animal. We know that damage to liver and other tissues can occur when too much juniper is consumed (Pritz et al. 1997). However, additional detrimental effects of juniper over-consumption are not well documented.
Management Practices That Might Enhance Juniper Consumption
Understanding why juniper is unpalatable to herbivores and how herbivores avoid the toxic effects of essential oils will lead to management practices that increase juniper consumption on rangelands. Greater consumption of juniper may be important to meet animal nutritional demands (especially during fall, winter, and during droughts) or enhance the use of goats in brush management.
Select Animals That Naturally Eat Large Amounts of Juniper
Some species and breeds of livestock eat more juniper than others (Straka 1993, Taylor et al.- chapter in this volume). Individuals within a breed also vary in their consumption of juniper. In a feeding trial with Spanish and Angora goats, certain individuals consistently ate more juniper than the herd average, while others consistently ate less than average (Pritz et al. 1997). As discussed earlier, these inter- and intraspecies differences result from differences in digestive morphology, detoxification capacities or physiological tolerances. Selecting the proper species, breed, or individuals within a herd or flock that naturally “like” and eat juniper could significancy alter the consumption of juniper in range settings.
Breed Animals with Desired Diet Characteristics
Many of the differences in diet preferences between individuals can be traced to inherited physiological, neurological, or morphological characteristics. Animal scientists and livestock producers have been quite successful at breeding animals with specific characteristics such as color, birth weight, hair quality, body morphology, horn shape, etc. It is very likely that we could breed goats that eat more than the average amount of juniper to other above-average goats to create a line of “juniper-eating” goats.
Exposure to Essential Oils Early In Life
There is growing evidence that previous dietary experiences influence the flavor preferences of animals and the ability of animals to digest, detoxify, and harvest certain plants (Launchbaugh 1996). Furthermore, experiences early in life often have a more lasting effect on consumption patterns than experiences later in life. Therefore, encouraging goats to eat juniper when they are young may enhance their ability to harvest and detoxify juniper when they get older.
With this concept in mind, we designed an experiment where young Angora and Spanish goats were given essential oils distilled from redberry juniper trees. The oils were given by capsules with a balling gun every other day for 30 days. We hypothesized, based on Brattsten 1983, that essential oils would be delivered to the liver and increase MFO detoxification activity. After the 30-day exposure period, we offered goats fresh juniper to eat. Interestingly, the goats that had received the essential oils ate similar (or slightly lower) amounts of juniper as the controls (Fig. 4; Pritz et al. 1997).
{kind=link}
There are several explanations for these results. Perhaps liver activity in goats is not induced by exposure to essential oils. Or, perhaps our doses of essential oils were too high or the goats were too young to respond positively. There was weak evidence that the doses of essential oils caused damage to the liver or other internal organs.
Nutritional and Pharmaceutical Supplements.
If increasing the consumption of juniper is desired, then starving animals onto the plants may not be the answer. Nutrient deprivation often decreases the rate of detoxification and increases an animal’s toxic response (Launchbaugh 1996). Supplementation of vitamins, mineral, amino acids and carbohydrates often enhances the ability of herbivores to detoxify or tolerate phytotoxins like essential oils (Boyd and Campbell 1983). Consequently, Taylor and associates (chapter in this volume) found that goats ate more juniper when given supplemental protein than when offered an energy supplement or no supplement at all. We have also tried to increase juniper consumption by supplementing animals with several compounds that aid in the detoxification of other chemically defended plants including: polyethylene glycol, monensin, and sodium bicarbonate. In each case, the supplemented animals ate no more than the control (unsupplemented animals). A more detailed understanding of detoxification pathways of essential oils will be necessary to develop a supplementation programs designed to entice rather than coerce livestock into eating juniper.
Conclusions
Converting juniper from a rangeland “weed” into livestock “feed” may be a lofty and unattainable goal. However, juniper is abundant and is relatively nutritious, especially compared to other range plants in the fall and winter, giving it several valuable forage characteristics. Unfortunately, there is a sturdy fence between the critter and “juniper forage”. The fence is the essential oils that juniper produces. Understanding how livestock detoxify these compounds and how the compounds affect juniper palatability will lead to management practices that will help the herbivore get over this allelochemical fence. These management practices are likely to include: selecting the proper breed or species of livestock, breeding animals with desired diet habits, giving them the proper early life experiences, and offering nutritional or pharmaceutical products to reduce the toxicity of the essential oils.
Literature Cited
Allison, M.J. 1978. The role of ruminal microbes in the metabolism of toxic constituents of plants. pp. 101-120. In: R.F. Keelir, K.R. VanKampen, and L.F. James (eds.), Effects of poisonous plants on livestock. Academic Press, New York, NY.
Boyd, J.N. and T.C. Campbell. 1983. Impacts of nutrition on detoxication. pp.287-306. In: J. Calwell and W.B. Jakoby (eds.), Biological basis for detoxication. Academic Press, New York, N.Y.
Brattsten, L.B. 1979. Biochemical defense mechanisms in herbivores against plant allelochemicals. pp. 200-270. In: B.A. Rosenthal and D.H. Janzen (eds.), Herbivores: Their interactions with secondary plant metabolites. Academic Press Inc., San Diego, Calif.
Brattsten, L.B. 1983. Cytocrome P-450 involvement in the interactions between plant terpenes and insect herbivores. pp. 173-195. In: P.A. Hedin (ed.), Plant resistance to insects. ACS Symposium Series #208. American Chemical Society, Washington D.C.
Cluff, L.K, B.L. Welch, J.C. Pederson, and J.D. Brotherson. 1982. Concentration of monoterpenoids in the rumen ingesta of wild mule deer. J. Range Manage. 35:192-194.
Garcia, J. and W.G. Hankins. 1975. The evolution of bitter and acquisition of toxiphobia. pp. 39-41. In: D. Denton and J. Coghlan (eds.), Olfaction and taste, Vol. 5. Academic Press, New York, NY.
Gershenzon, J. and R. Croteau. 1991. Terpenoids. pp. 195-219. In: G.A. Rosenthal and M.R. Berenbaum (eds.), Herbivores: Their interactions with secondary plant metabolites. Academic Press Inc., San Diego, Calif.
Hofmann, R.R. 1988. Anatomy of the gastro-intestinal tract. pp.14-43. In: D.C. Church (ed.), The ruminant animal: Digestive physiology and nutrition. Waveland Press, Inc, Prospect Heights, Ill.
Huston, J.E., B.S. Rector, L.B. Merrill and B.S. Engdahl. 1981. Nutritional value of range plants in the Edwards Plateau region of Texas. Texas A&M Exp. Sta. Bull. #B-1357. Texas A&M University, College Station, Tex.
Launchbaugh, K.L. 1996. Biochemical aspects of grazing behavior. pp. 159-184. In: J. Hodgson and A.W. Illius. The ecology and management of grazing systems. CAB International, Wallingford, U.K.
Launchbaugh, K.L. and F.D. Provenza. 1994. The effect of flavor concentration and toxin dose on the formation and generalization of flavor aversions in lambs. J. Anim. Sci. 72:10-13.
Laycock, W.A., J.A. Young, and D.N. Ueckert. 1988. Ecological status of poisonous plant on rangelands. pp. 27-42. In: L.F. James, M.H. Ralphs, and D.B. Nielson (eds.), The ecology and economic impacts of poisonous plants on livestock production. Westview Press, Boulder, Colo.
Lindroth, R.L. 1989. Mammalian herbivore-plant interactions. pp. 163-205. In: W.G. Abrahamson (ed.), Plant-animal interactions. McGraw-Hill Book Co., New York, NY.
Molyneux, R.J. and M.H. Ralphs. 1992. Plant toxins and palatability to herbivores. J. Range Manage. 45:13-18.
Nagy, J.G., H.W. Steinhoff, and G.M.Ward. 1964. Effects of essential oils of sagebursh on deer rumen microbial function. J. Wildl. Manag. 28:785-790.
Oh, H.K., T. Sakai, M.B. Jones, and W.M. Longhurst. 1967. Effects of various essential oils isolated from douglas fir needles upon sheep and deer microbial activity. Applied Microbiol. 15:777-7784.
Pritz, R.K., K.L. Launchbaugh, and C.A. Taylor. 1997. Effects of breed and dietary experience on juniper consumption by goats. J. Range Manage. 50: (in press).
Provenza, F.D. 1995. Postingestive feedback as an elementary determinant of food preference and intake in ruminants. J. Range Manage. 48:2-17.
Riddle, R.R., C.A. Taylor, M.M. Kothmann, and J.E. Huston. 1996. Volatile oil contents of ashe and redberry juniper and its relationship to preference by Angora and Spanish goats. J. Range Manage. 49:35-41.
Schwartz, C.C., W.L. Regelin, and J.G. Nagy. 1980. Deer preferences for juniper forage and volatile oil treated foods. J. Wildl. Manage. 44:114-120.
Straka, E.J. 1993. Preferences for redberry and blueberry juniper exhibited by cattle, sheep and goats. M.S. Thesis. Texas A&M Univ., College Station, TX.
Taylor, C.A., J.E. Huston, Jr., N.E. Garza, T.D. Brooks and R.A. Moen. 1994. Use of juniper as a supplemental feed limiter. p. 31-33. In: Sheep and Goat, Wool and Mohair Report. PR-5222. Texas A&M Experiment Station, College Station, TX.
Comments: Allan McGinty, Professor and Extension Wildlife Specialist