F. E. Smeins and S. D. Fuhlendorf
Distribution and Abundance
Ashe juniper (Juniperus ashei Buchholz)(1) is found from southern Missouri, through Arkansas into Oklahoma, across central Texas and into Mexico and Guatemala. It occurs primarily on calcareous, shallow, rocky soils derived from limestone and dolomitic parent materials, although it can also grow on sandy, granitically derived soils (Vines 1960). In Texas, it is found in greatest abundance on the eastern and southern portions of the Edwards Plateau, but also extends into the Central Mineral Region, Grand Prairie, Cross Timbers and North Central Prairies Ecoregions (Texas State Soil and Water Conservation Board 1991).
The exact distribution and abundance of Ashe juniper on the Edwards Plateau at the time of European settlement may never be known with any degree of accuracy. Early observers, recorded very dense and nearly closed-canopy “cedar brakes” in some areas, particularly on the canyon slopes and on rocky, sloping land of the Balcones canyonlands over Glen Rose limestone substrates. On the other hand, Ashe juniper was apparently of minimal density in many areas, particularly on the divide portions of the central and northwestern Plateau (Smeins 1980, Weniger 1984, Smeins et al. this volume).
In spite of lack of resolution about its original distribution and abundance, since European settlement this species has generally increased in abundance and habitat range with fluctuations based on periodic naturally and man-caused fires or efforts to reduce it or harvest it for various uses and commercial products (Bray 1904, Foster 1917, Smeins et al., this volume, Garriga this volume). Since European settlement large areas of cedar brakes have been harvested for fence posts and other purposes (Cartwright, 1966), but following harvest Ashe juniper tends to rapidly reestablish itself (Huss 1954, Weniger 1984). Thus, reestablishment in previously occupied areas, as well as spread into new sites is an ongoing process particularily in the absence of periodic fires.
The exact cause of increase in Ashe juniper is difficult to determine. Overgrazing by livestock, which reduces herbaceous plant competition for establishing juniper seedlings is frequently considered a factor. Purposeful or inadvertent prevention of naturally occurring fires, along with reduced fuel loads due to grazing, have also had an influence. Since this species does not sprout from the base when it is topkilled, it is readily controlled by fire if sufficient fuel is available and if the plants are burned at less than mature size. Wink and Wright (1973) found that 99% of Ashe juniper plants less than 1.8 m tall(2) were killed with a fine fuel (grass) load of 1500 kg/ha or more. As the trees become larger, the denuded area beneath their canopy expands and it becomes increasingly difficult to ignite the plants or to move the fire to the fire-sensitive central stem (Dalrymple 1969). Periodic droughts, particularly when associated with overgrazing, may also play a role in their spread. The weakened, open herbaceous cover could be less resistant to juniper invasion following droughts when favorable rainfall conditions return. On the other hand, long-term droughts, such as the one of the 1950’s, can cause extensive mortality of large, mature, Ashe juniper, although, smaller size classes apparently survive droughts better than associated species such as live oak (Quercus virginiana Mill.) and shin oak (Q. pungens Liebm. var. vaseyana (Buckl.) C.H. Muller) (Merrill and Young 1959). Historical soil erosion due to overgrazing, droughts and other factors may increase the heterogeneity of sites making them more susceptible to woody plant invasion. Documented increase in atmospheric carbon dioxide since the 1850’s, which may favor growth conditions for juniper and other woody species, compared to the warm season grasses, may also be a factor (Emanuel et al. 1985). As with most ecological phenomena, the increase of Ashe juniper is most probably an interaction of all these factors (Smeins et al., this volume).
Ashe juniper significantly alters the composition and structure of rangeland plant communities. Increasing density of Ashe juniper reduces grazeable areas for livestock and many species of wildlife because of its low forage value. It significantly reduces the production and diversity of associated plant species (Blomquist 1990, Fuhlendorf 1992, Yager 1993, Fuhlendorf and Smeins 1997). Additionally, growth of Ashe juniper in the immediate vicinity of more desirable woody species such as live oak, not only denies browsing animals access to these species but may in time cause mortality of the desired species (Rykiel and Cook 1988). Ashe juniper does, however, serve as cover and food for many wildlife species. Deer and domestic livestock may consume large quantities of berries, particularly when other forages are of limited availability or quality. Bark from certain size classes of Ashe juniper is apparently essential as nesting material for the endangered golden-cheeked warbler (Dendroica chrysoparia) (Kroll 1980). Ashe juniper may influence the hydrology of an area when it grows in abundance. Alternatively, it may limit soil erosion on many sites including steep slopes of canyons where other vegetation cover cannot grow or is sparse (Wright et al. 1975, Marsh and Marsh 1993).
To better understand the variation in abundance of this species, its influence on ecosystem structure and function and to more effectively develop strategies for its management, a detailed understanding of its biology and ecology is essential. This chapter will summarize our knowledge of Ashe juniper in terms of its biogeography, taxonomy, life history characteristics, and relationships with other plants and animals in the ecosystem.
Growth Habit and Taxonomy
Ashe juniper(3) is an evergreen, dioecious (separate male and female plants) shrub or small tree that is usually less than 6 m in height at maturity. It is non-sprouting, typically multistemmed, and has a broadly globular growth form that usually branches near the base. Its stems are often fluted and twisted and, as they age, they develop a gray to reddish brown bark that shreds into shaggy longitudinal strips. Its wood is dense with a pale-brown heartwood and a broad zone of nearly white sapwood. Leaves on mature branches are scalelike and occur in pairs or sometimes threes which are closely appressed, triangular to ovate in shape, and average 2 mm long and 1 mm wide. Leaves on young shoots and seedlings are spiny and awl-like and may be up to 12 mm long. Small, inconspicuous flowers are located terminally on branches. Staminate flowers are 4 mm long with 12 to 18 stamens; they are brown in color. The green pistillate flowers are up to 3 mm long and are formed by cohesion of enlarged fleshy scales to form a berrylike cone. As the one-seeded (sometimes two) berry matures, it takes on a deep blue color and varies in size from 6.5 to 8mm in length and 5.5 to 7mm in width (Chavez-Ramirez and Slack 1994). Seeds are 2 mm long with a thick woody seed coat (Correll and Johnston 1970).
The origin of Ashe juniper is not known although most believe it to be part of the southwestern Madrean Evergreen Woodland that occurs in Mexico and extends into New Mexico and the Southwestern U.S. (Brown 1982). It overlaps in geographic distribution with Eastern Red Cedar (Juniperus virginiana L.) to the east and north and with Redberry Juniper (Juniperus pinchotii Sudw.) to the west and northwest. Based upon anatomical and morphological features it has been suggested that Ashe juniper may form hybrids with both of these species where they come into contact with one another (Hall et al. 1961, Gehlbach 1988). Some authorities feel that Ashe juniper is closely related to one-seeded juniper (Juniperus monosperma (Engelm.) Sarg.), which occurs from western Texas and Oklahoma through New Mexico to Utah, Colorado and Wyoming, and some would consider one-seeded juniper to be only a variety of Ashe juniper (Vines 1960). Chemical analysis of the foliage of Ashe and other junipers suggests that it is chemically distinct from other species, and no chemical evidence exists of hybridization between Ashe juniper and other species that overlap its geographic range (Rudloff 1968, Adams and Turner 1970). The final judgment of its genetic, biogeographic, and taxonomic status has yet to be confirmed.
LIFE HISTORY
Introduction
Life history refers to the set of adaptations of an organism that more or less directly influence its survival and reproductive success. It includes reproductive rate and output, age at first reproduction, seed germination, longevity and dispersal, growth rate and pattern as well as many other characteristics of the organism. For Ashe juniper information is available for some of these parameters while for others they remain unknown or are currently being analyzed.
Flowering and Seed Production
Flowers are typically produced between December and February with pollen release and pollenation occuring during this time. Cone (fruit, berry) development and maturation occurs from then until November, and fruit ripening and dispersal between November and April. Observations indicate that Ashe juniper is capable of producing abundant berry and seed crops. Trees begin to produce fruits at about 1 to 1.5 m in height (approximately 10 to 20 years old). Fruit and seed production varies greatly from tree to tree within a year and for the same tree across years (Table 1). A given tree may produce abundant fruit crops in successive years, if environmental conditions are favorable, which suggests that environmental rather than genetic limitation controls berry (seed) crops. Large trees may produce between 100,000 to 250,000 berries per tree. Smaller trees produce proportionately fewer berries (Table 1).
Weather (precipitation) appears to be a controlling factor in berry production. For example, several large trees on the Sonora Research Station in 1989-90 (wet year) produced 50,000 to 200,000 berries per tree, while in 1990-91 (dry year) they produced fewer than 1,500 berries and in 1991-92 (wet year) they all produced in excess of 100,000 berries (Table 1) (Chavez-Ramirez 1992). A plot with 18 fruit bearing trees in 1989-90 (wet) had only 4 bearing fruit in 1990-91 (dry). Reinecke (1996) also found mature trees that had prolific seed production in 1993 (wet) had limited production in 1994 (dry).
Table 1. Fruit production of four large Ashe juniper trees over a three year period. (From Chavez-Ramirez 1992)
| Fruit Numbers Per Tree | ||||
| Tree | CanopyArea(m2) | 1989-90(wet) | 1990-91(dry) | 1991-92(wet) |
| 1 | 16 | 57,500 | 1,500 | no data |
| 2 | 28 | 91,000 | 0 | 105,000 |
| 3 | 33 | 196,000 | 100 | 226,900 |
| 4 | 76 | 154,600 | 90 | 198,600 |
Seed Rain and Dispersal
Seeds of Ashe juniper are relatively heavy and hence those that fall from the parent tree tend to remain within or very near the canopy area. If not removed by predators, most seeds fall from the tree within days after ripening. Near Uvalde, Texas, 40 trees were marked that had abundant seed production. Even though there were abundant birds and mammals (seed foragers) present, few if any of the berries were consumed. Most fell directly under or near the parent tree (Owens and Schliesing 1995). Perhaps in some years berry production exceeds a threshold and is so high that even a great number of seed foragers may miss individual or entire stands of trees with abundant berry production. Holthuijzen and Sharik (1984) found most seeds of the similar Eastern Red Cedar to fall beneath the canopy and few seeds were found beyond 5 m of the parent plant. On relatively level sites on the Sonora Research Station no Ashe juniper seeds were found in soil samples over 5 m from parent plant canopies (Kinucan and Smeins 1992). If the tree occurs on a slope or an area that has periodic overland flow of water during rainfall events, many seeds can be carried by gravity or water and deposited some distance away (Huss 1954). When deposited, they are often in sites with a collection of well-watered debris and litter that may favor germination and seedling establishment.
Berry (seed) foragers may remove nearly all berries from a given tree within a few hours or days of ripening. Primary agents of long distance dispersal are birds and mammals. Berries are readily consumed by nearly all mammals, including all classes of livestock, many bird species and some insects. Raccoons (Procyon lotor)(4) and ringtail cats (Bassariscus astutus) as well as gray foxes (Urocyon cinereoargenteus), coyotes (Canis latrans) and jackrabbits (Lepus californicus) consume the berries. American robins (Turdus migratorus) and cedar waxwings (Bombycilla cedorum) are important bird foragers primarily because of their very high winter populations in the Ashe juniper region (Chavez-Ramirez and Slack 1993, 1994).
Not only do all these animal species consume the seeds but large numbers of ingested seed avoid destruction by mastication or digestive processes and pass intact through the animal and exit with the feces. As a result they may be deposited some distance from the parent plant (Chavez-Ramirez 1992, Chavez-Ramirez and Slack 1993). Robins are perhaps more effective dispersal agents than waxwings, due to their different post-foraging behavior (Chavez-Ramirez and Slack 1994). Robins have a much greater range of movement away from parent trees following ingestion of juniper berries and they tend to individually visit a wider variety of perch sites, particularly within grassland areas.
Berries are harvested by animal foragers and disperers when the berries are ripe (fully hydrated). It was speculated that seeds would be at their peak of germination potential at this time. Samples of berries were collected from a series of mature Ashe juniper trees on the Sonora Reasearch Station periodically from mid-October 1993 to mid-February 1994 and again from October to December 1994. Time of peak germination and percent germination of cold-treated seeds varied dramatically between trees (see seed germination section). Some trees never produced seeds that germinated while other trees had a high of 55% germination on a given date. Each tree had a unique germination pattern. Some were highest in the fall, others in mid-winter and other in late-winter. Highest germination, for any tree in many cases, corresponded to when the berries were ripe (Reinecke 1996). Thus, it appears that synchronous maturation of seeds occurs within a given tree which coincides with berry maturation and palatability. Seeds at other trees mature and berries ripen at other times over as much as a 4 to 5 month period. This combination of events i.e. synchronous maturation within a tree and asynchronous maturation between trees, effectively favors seed consumption and dispersal by animals and retains dispersal agents, such as robins and waxwings in the area for a long period of time. It also provides an important forage component for livestock and white-tailed deer during a time of low forage availability.
Seed Bank
The number of seeds in the leaf litter and top 8 cm of mineral soil beneath 18 large female Ashe juniper trees was determined on the Texas A&M Research Station at Sonora. The number of intact seeds was greatest in the litter and varied from 11,000 to 23,000 seeds/m2. The mineral soil varied from 250 to 650 seeds/m2. Overall average was 16,600 seeds/m2 in the litter and top 8 cm of the soil (Blomquist 1990). These seeds were the result of seed rain from several previous seasons. Tests were conducted to test potential viability. Although viability was only <0.3%, the large number of seeds in the seed bank still would result in nearly 50 viable seeds/m2. Most of the viable seeds were considered to be seeds produced in the most recent seed crop. Owens and Schliesing (1995) found between 1197 and 1436 seeds/m2 in the leaf litter and 275 to 366 seed m-2 in the soil on a site nor Uvalde. High variation across studies would be expected because of variable seed production from year to year.
Seed Germination
Berries were collected from several trees on the Texas A&M Research Station during the fall of 1987 and subjected to treatments that may naturally occur. Seeds were germinated after receiving the following treatments: (1) no treatment; (2) berries fed to goats, raccoons and robins and seeds collected from the feces; (3) berries mechanically removed from the seeds. Subsamples of all of the above were cold stratified at 2o C for 2 months and their germination compared to non-cold treated seeds.
Digestive scarification did not significantly influence seed germination over mechanical removal of berries. Mean germination across all treatments was 29%. Cold stratification significantly increased average germination to 45%. With the berry removed, germination equalled 32%; while with the berry intact it was only 0.02%. Seeds from individual trees varied from 30 to 50% germination with the fruit removed. The fruit appears to act as a physical or chemical inhibitor to germination. The advantage of seeds passing through the digestive tract of an animal may be more influenced by removal of the berry rather than any physical or chemical scarification of the seeds due to the digestive process.
It appears that favorable precipation during the late spring-early summer period is related to the degree of germination of seeds produced in that year. Total annual rainfall or that of other months or seasons was not strongly correlated with level of germination (Reinecke 1996). Significant tree to tree variation occurred in amount (from 0 to up to 55% peak germination) and timing of germination (some peaked as early as mid-October and others as late as early January). Cold-stratification of seeds with the fruit removed increased overall germination from 0 (non-stratified) to 15% (cold-stratified) across all trees and sample dates. Presence of the berry resulted in no germination of stratified or non-stratified seeds. Laboratory storage of seeds for 6 and 24 months did not significantly alter germination responses.
Seed Longevity
Seeds were harvested from trees on the Sonora Research Station during the fall of 1987. They had an average cold-treated germination percentage of 45% immediately after collection. They were placed in nylon bags and either positioned on the soil surface or buried 1 to 2 cm below the soil surface. Samples were collected every 6 months for two years and germination tests conducted. By the end of 18 months, germination had dropped to less than 1% regardless of being on the soil surface or buried in the seed bank. This suggests that seed persistence is rather limited under field conditions, although 1% of the hundreds of thousands of seeds produced in a stand of mature trees would still be a significant number for potential recruitment into the population.
On a site near Uvalde germination of seeds from the seed bank was 0%, while newly collected seeds of the year was 5% (Owens and Schliesing 1995). These results are similar to those found by Holthuijzen and Sharik (1984) for Eastern Red Cedar where <5% of seeds remained viable after 400 days. Reinecke (1996) found, however, that seeds kept in the laboratory for up to 24 months retained the same levels of germination as immediate post-collection germination trials. The reason for the contrasting results is uncertain, however, it is believed, and inspection of field placed seeds agrees, that in the field dessication, bacterial and other degrading factors seem to destroy the seeds after 18 months.
Seedling Distribution and Survival
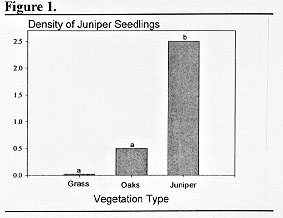
Samples were taken from under juniper trees or oak mottes and out in open grassland from pastures with different grazing histories across the Sonora Research Station to determine the numbers and distribution of seedlings (Blomquist 1990). The greatest density of seedings (2.5/m2) occurs under female juniper trees, whereas density under oaks or in open areas is substantially less (Figure 1). However, there could be great turnover of these seedlings under juniper canopies, that is, they may establish only to die during the following year and be replaced by seedlings from the next seed crop. Observation would suggest that some of this phenomenon may occur but many seedlings under parent trees appear to have survived for several years. They have thick basal stems with several annual rings, their leaves are of the mature form, and some are up to 0.5 m in height. Thus as juniper trees become larger they serve as a large reservoir of seedlings either due to protection from herbivores or as favorable establishment sites.
{kind=link}
Death of the large overstory tree can release many seedlings that may grow rapidly due to increased light, moisture and nutrient availability. Where one large tree occurred a cluster of small plants can develop. Thus, removal of large trees by mechanical methods not followed by burning or other practices may result in rapid replacement of the removed plants.
Another study on the Sonora Station evaluated numbers of Ashe juniper seedlings and saplings under juniper and oak canopies and adjacent grass patches in four longterm grazing treatments (Reinecke 1996). Greatest densities were under female junipers followed by male trees and oak mottes. Grass patches had very low densities. Female trees in pastures with a previous history, but not current heavy browsing by goats, had the highest seedling densities. This is perhaps a result of the still existing browse line on these previously heavily browsed plants which now provides a better light environment for seedling survival. This treatment also had the lowest juniper density and cover.
It would appear that seedling establishment is possible across a great variety of soil/vegetation conditions. They may more readily establish in heterogeneous, rocky, relatively, open sites, however, they can and do establish on relatively deep soils with a continuous cover of vigorous herbaceous plants. In the latter case, increase in size and numbers may be slowed by herbaceous competition but invasion of juniper is ultimately possible. Ability to establish under the dense canopy of parent juniper and other woody species suggests that high light intensity is not essential for their germination or establishment but may favor their survival.
Van Auken (1993) evaluated closed juniper woodlands and adjacent grassland opening on eastern portions of the Edwards Plateau in Bandera County. He found high densities of Ashe juniper seedlings in heavily grazed grasslands and concluded that the low grass biomass due to grazing reduced interference for establishment of juniper seedlings and also reduced fire potential. He inidcated low densities under mature canopies of the closed woodland perhaps due to low light intensities. Comparison of this area, on quite different soils and climatic conditions, to the more westerly location of the Sonora Research Station, must be made with caution since the different environments, could produce contrasting results. Stands at Sonora never reach the crown closure of mature stands on the eastern Plateau. Hence, his observation that mature stands exhibit declining seedling densities could be a general but yet undocumented phenomenon. Most stands are regrowth from former thinning and clearing and most have, perhaps, not reached a mature stage of growth and corresponding decline in seedling survival.
A recent investigation near Comfort, Texas suggests that grazing and the presence of grass vegetation, may influence seedling survival different from the conclusions of Van Auken (1993). Four treatments, ungrazed-high grass cover, ungrazed-low grass cover, grazed-high grass cover and grazed low-grass cover were evaluated (Reinecke 1996). Of 1696 new seedlings marked during the winter of 1995, there was on overall 45% survival across all treatments after nearly 500 days the survivorship in each treatment was: ungrazed/ high grass cover-60%; ungrazed/low grass cover-49%; grazed/high grass cover-40%; grazed/low grass cover-31% (Figure 2).
{kind=link}
These results suggest that moderately stocked cattle, white-tailed deer, rabbits, armadillos, etc. do reduce the number of seedlings through consumption and trampling. Herbivore use and selection of juniper seedlings is discussed in more detail by Taylor et al. (this volume). Also, high grass cover may enhance seedling survivorship by providing protection from herbivores and a favorable microenvironment for growth and development. Variable conclusions from seedling establishment studies suggest that germination and survival of juniper may be episodic where some years low grass cover may enhance establishment while other years, high cover may be more critical.
Growth of Individual Plants
Once established, rate of growth and development determines the influence of the juniper trees on the rest of the community. In 1949, nearly all junipers were hand-removed from the Texas A&M Research Station. Aerial photographs from 1955, 1966, 1973 and 1985 were obtained and 14 trees that could be identified on photographs from 1955 were followed through each photograph to assess their individual rate of canopy increase (Blomquist 1990). The average canopy showed a steadily increasing growth rate over the 30 year period: 0.58 m2/yr from 1955 to 1966, 1.40 m2/yr from 1966 to 1973, and 1.94 m2/yr from 1973 to 1985 (Figure 3). Great variation existed in individual growth patterns. Some plants exhibited little, if any, growth during the first 10 to 20 years and then grew rapidly. For example, one tree expressed little growth from 1955 to 1973 but then increased in size by 42 m2 from 1973 and 1985, a rate of 3.50 m2/yr. Others grew rapidly at first and then greatly reduced their growth rate. Differences appear to be related to different site conditions. Although first production of seeds occurs at approximately 1 to 1.5 m in height or 10 to 20 years, as their age and size increase their quantity of seed production increases.
{kind=link}
COMMUNITY RELATIONSHIPS
General Composition-Edwards Plateau
Ashe juniper has been a component of the plant communities of the Edwards Plateau for thousands of years (Bryant and Shafer 1977, Smeins et al. this volume). The general geographic range of the species has probably not changed greatly since the time of settlement. It has, however, increased in density in communities where it has always occurred, and in some instances spread into habitats where it previously was absent or of limited abundance (Smeins 1980, Riskind and Diamond 1988, Van Auken 1988). Riskind and Diamond (1988) provide an excellent general overview of the plant communities and environments of the Edwards Plateau and indicate, along with others such as Gehlbach (1988) and Van Auken (1988), that Ashe juniper can successfully grow on nearly all geology/soil sites. Hence, given sufficient time without fire or heavy use by browsers and adequate dispersal, Ashe juniper can probably become a dominant plant on nearly all sites on the Edwards Plateau, as well as many adjacent ecoregions.
Composition – Texas A&M University Research Station
Canopy cover and density of woody plants was determined for pastures on the Texas A&M Research Station that have had 3 different grazing histories since 1948 (Tables 2 and 3). Although, most juniper had been hand cut from the Station in 1948, canopy cover was significantly different across the three grazing histories in 1984. The historically (1949-1969) heavily browsed (goat) pastures had 15% total canopy cover which was less than one-half the cover of the ungrazed pastures (exclosure), while light browsed (Merrill deferred rotation) pastures had intermediate values. Percent woody cover of live oak varied from 3.3 to 7.5%. Shin oak had only 2.1% cover in heavily browsed pastures, but 14.1% in ungrazed pastures. Ashe juniper varied from 4.4 to 11.1% with highest values in ungrazed pastures (Table 2). Since no fires have occurred in these pastures since 1948, it appears that long-term grazing does not contribute to the increase in woody plants and may actually inhibit it.
Eighteen woody species were encountered in ungrazed pastures while light and heavy browse pastures had 12 and 10 species, respectively. Thirty-five percent of total cover was contributed by other woody species in the heavy browse pasture and the majority of that was made up of Texas persimmon (Diosporous texana Scheele) and algerita (Mahonia trifoliata Moric. Fedda). Elbowbush (Forestiera pubescens Nutt.), hackberry (Celtis laevigata Willd.) and skunkbush sumac (Rhus aromatic Ait), which are preferred browse species, were absent from light and heavy browse pastures, while in ungrazed pastures the latter two species made up the majority of the composition of other woody species.
Ashe juniper size and density was significantly affected by grazing history (Table 3). Total density of heavily browsed pastures was 276 plants/ha which was approximately 2.5 times less than light and no browsed pastures. For all height classes heavily browsed pastures had fewer plants than other grazing histories except for the smallest size classes where no significant differences were recorded (Table 3). Heavy browsing by goats from 1948 to 1969 has continued to dramatically limit the invasion and growth of juniper until recent years. There are fewer large berry producing trees in these pastures which further reduces the opportunity for new recruitment.
Table 2. Percent cover of woody species for different grazing histories. Different letters indicate significant differences ( 0.05) between grazing histories (from Station records).
| Percent Cover | |||
| NoGraze | LightBrowse | HeavyBrowse | |
| Live Oak | 7.4 a | 7.5 a | 3.3 a |
| Shin Oak | 14.1 b | 8.6 b | 2.1 a |
| Ashe juniper | 11.1 b | 6.3 ab | 4.4 a |
| Other species | 4.4 a | 3.6 a | 5.2 a |
| Total | 37.0 c | 26.0 b | 15.0 a |
Table 3. Stem densities of Ashe juniper by canopy diameter size class for three grazing histories, Different letters indicate significant differences (< 0.05) between grazing histories (from station records).
| Grazing History | |||
| CanopyDia. (m) | NoGraze | LightBrowse | HeavyBrowse |
| 0.0-0.25 | 170 a | 143 a | 145 a |
| 0.25-1.0 | 161 a | 176 a | 69 a |
| 1.0-2.0 | 257 a | 205 a | 34 b |
| 2.0-3.0 | 107 a | 117 a | 7 b |
| 3.0-4.0 | 26 a | 14 ab | 4 b |
| >4.0 | 47 a | 24 b | 17 c |
| Total | 769 a | 679 a | 276 b |
Longterm Change in Woody Plant Composition
Photographs from the early 1900’s indicate that juniper was a small portion of the woody vegetation of the Texas A&M Research Station. By the 1940’s it had become a significant portion of the plant community. In 1949 most junipers were removed from the station by hand cutting. Following the drought of the 1950’s Merrill and Young (1959) evaluated the impact of the drought on woody species mortality and survival. The pre-drought composition of woody plant cover was live oak 64%, shin oak 26%, Texas persimmon 4% and juniper 1%. The remaining 5% was composed of several species including hackberry, algerita, elbowbush and catclaw acacia (Acacia greggii Gray). Live oak suffered a 54% mortality and shin oak 30%. At the end of the drought the composition had shifted slightly to 52% canopy cover for live oak, 33% for shin oak, 8% for Texas persimmon and 2% for juniper. Large junipers suffered 90% mortality while plants less than 2 m in height experienced no death loss. Overall there was a 56% reduction in canopy cover of all woody plants. Thus, extended droughts can significantly influence the abundance and composition of the woody vegetation, and appears to favor increases in juniper.
Interpretation of aerial photographs from 1955, 1966, 1973 and 1985 provide an assessment of longterm change in Ashe juniper numbers and canopy cover for selected sites on the Texas A&M Research Station at Sonora. The number of canopies increased from approximately 15/ha in 1955 to 82/ha in 1985. The majority of this increase occurred between 1973 and 1985. Percent juniper cover increased from less than 1% in 1955 to over 12% in 1985 (Table 4). The majority of this increase also occurred from 1973 to 1985 which agrees with the period of time of greatest increase in growth of individual trees described above. Fuhlendorf (1992) found similar levels of increase in Ashe juniper using interpretation of aerial photographs on an entire grazing exclosure on the Station. Smeins and Merrill (1988) documented an overall increase in total woody plant cover of 12 to 32% from 1949 to 1983 across several grazing regimes. Juniper increased from 5% of the composition to 35% over that time period which accounted for the majority of the total increase.
Table 4. Density (no/ha) and percent cover of juniper canopies at 4 dates over a 30 year period. Different letters denote significant differences (p < 0.05) (From Blomquist 1990).
| Year | ||||
| 1955 | 1966 | 1973 | 1985 | |
| Density of Canopies | 14 a | 25 a | 30 a | 82 b |
| Percent Cover | 1 a | 3 a | 4 b | 12 c |
Samples from several pastures across the Sonora Station collected in 1993 showed all pastures, regardless of grazing regime or stocking rate, to have about the same level of woody canopy cover (40%) and Ashe juniper contributed >50% of the composition. The only exception was again the historically heavily goat browsed pastures that had lower total cover 17% with about the same proportion for juniper.
The ability of juniper to increase in canopy cover is either the result of increase in size of existing plants or recruitment of new plants. Minimal increase in canopy cover from 1955 to 1973 suggests that slow growth rate of the small plants and limited recruitment kept total canopy cover low. However, by 1973 sufficient plants had reached adequate size to rapidly increase their growth rate and by seed production which resulted in increased recruitment potential. Collectively this resulted in rapid canopy cover increase from 1973 to 1985 that continues today.
Age/Size Relationships
Age/size relationship were determined for 75 trees varying in height from <0.5 m to over 4 m collected from several different pastures on the Sonora Research Station in 1991 (Fuhlendorf 1992). Trees were cut at ground level with a chainsaw and cross-sections were removed from the base to determine annual ring counts. Ring counts, as estimates of age, were then related to plant height and canopy diameter for each tree to determine age/size relationships (Table 5). The trees varied in age from 11 to 77 years.
An attempt was made to remove all junipers from the study site in 1949 by hand cutting, thus the maximum age of trees should be about 42 years old. However, field vegetation surveys (Merrill and Young 1959) and photographs of the station in the early 1950’s indicate the presence of Ashe juniper trees, some of substantial size, which suggests that all plants were not removed in 1948. Other woody species were not removed, and the high densities of these species could explain the failure to remove all junipers, since the other species may have provided escape cover for junipers.
Analysis of trees less than 40 years old showed those with a long history of heavy browsing by goats were significantly smaller in size at a given age than those from pastures with a history of light browsing (Table 5). Trees between 10 and 40 years of age had generally one-half the canopy diameter and heights in heavily browsed pastures compared to pastures with a history of light browsing. As trees became larger, the influence of utilization of goats decreased and those trees over 40 years of age showed no differences between grazing histories. Pruning of the juniper by the goats apparently is the cause of these differences. Therefore, heavy browsing by goats reduces the canopy growth rate of individual trees, as well as maintaining lower densities and cover within a pasture.
Table 5. Average canopy diameters (m) and height (m) of Ashe juniper for 4 age classes on the Texas A&M Research Station, 1991. Different letters indicate significant differences between grazing histories. (from Fuhlendorf 1992)
| Pastures |
| AgeClass (yrs) | HeavyBrowse | LightBrowse | |
| Canopy Diameter | 10-20 | .66 a | 1.18 b |
| 21-30 | 1.43 a | 2.43 b | |
| 31-40 | 1.41 a | 2.69 a | |
| >40 | 5.56 a | 5.41 a | |
| Height | 10-20 | .82 a | 1.52 b |
| 21-30 | 1.48 a | 2.35 b | |
| 31-40 | 1.51 a | 2.67 a | |
| >40 | 4.46 a | 4.37 a |
Overstory/Understory Relationships
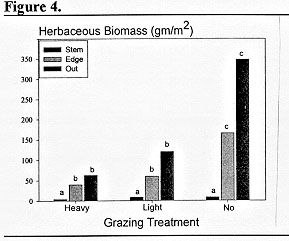
Ashe juniper plants from small saplings to large trees were selected within ungrazed exclosures, moderate-deferred rotation pastures and heavily-continuously grazed pastures to determine the influence of past grazing history and juniper canopy on herbaceous production (Blomquist 1990). Samples were collected from near the stem base, the edge of the canopy and in open areas away from the canopy (Figure 4). The zone from the stem base to near the canopy edge typically has little herbaceous production or it is very low, generally less than 5 gm/m2. Hence, as the juniper plant gets larger, more and more ground is lost to production of herbaceous forage and the forage produced is often inaccessible to grazing animals because of the dense, low growing juniper foliage. The dense shade of the juniper canopy as well as the deep accumulation of juniper leaf litter under larger trees appear to be the primary constraints to herbaceous production (Fuhlendorf and Smeins 1997). The species diversity of herbaceous species is also greatly reduced by increased canopy cover and litter accumulation of juniper(Fuhlendorf 1992, Yager 1993).
{kind=link}
There is little evidence that the accumulated litter of Ashe juniper in anyway alters the chemical nature of the soil as it relates to growth and development of other plants (Yager 1993). Soil chemical and physical properties are in fact “improved” by the presence of the juniper litter (Marshall 1995). No allelopathic effects have been shown to be produced by the litter. The major impact of the litter seems to be its physical presence and its alteration of hydrologic properties of the area under the canopy. The thick litter layer is a difficult physical medium for seeds of other species to germinate and grow in mainly because moisture either runs off (due to the hydrophobic nature of the litter) or it dries out very quickly after a precipitation event, which prevents seedling growth from reaching the mineral soil (Yager 1993).
When heavy goat browsing has been a factor in a pasture, a browse line is created on the juniper which alters their influence on herbaceous production and cover as compared to unbrowsed plants (Fuhlendorf 1992). The understory of browsed plants has more light penetration, less litter accumulation and generally more herbaceous production and species diversity than unbrowsed plants. A fairly dense cover of cool season grasses, such as Texas wintergrass (Stipa leucotricha Trin. & Rupr.), is often prevalent under browsed canopies partially reducing the influence of Ashe juniper on herbaceous production and diversity. Figure 5 is a schematic side view diagram which illustrates the interaction of the browse line, litter depth and understory herbaceous foliar cover for a tree with a history of heavy browsing verses one that has a history of no browsing (Fuhlendorf 1992).
{kind=link}
Successional/Disturbance Relationships
Observation and most studies indicated that Ashe juniper has great potential to re-establish
itself on sites from which it has been removed and in the absence of fire to spread to sites that it historically did not occupy. It would appear that much of the Edwards Plateau could succeed to nearly closed canopy stands in the absence of periodic disturbances, such as fire. Of course, grazing regimes, fire regimes, weather variation and other factors interact to influence the rate and pattern of these changes. Longterm models of grassland/woodland vegetation dynamics are presented by Fuhlendorf (this volume). Transformation from a grassland to a woodland dominated landscape occurs when fires are absent, regardless of grazing treatment. This change is associated with a threshold, where change is slow and reversible initially but after 25-50 years (depending upon grazing and weather) become rapid and non-reversible with typical management fires. Emergent processes related to these changes and effects on hydrology, wildlife, livestock and other resources and uses are discussed in other chapters of this volume (Thurow, Owens and Ansley, Rollins and Armstrong, Kothman and others).
Summary And Implications
1. Ashe juniper is a small, non-sprouting, evergreen, dioecious tree that occurs primarily on limestone substrates from Central America to Missouri with its center of abundance within the Hill Country of Texas.
2. Ashe juniper has always been abundant on portions of the Edwards Plateau but it is generally agreed that its habitat range and overall abundance has increased since European settlement/or at least during the past 50 years
3. Elimination of naturally occurring fires, overgrazing, soil erosion, droughts, increase in atmospheric carbon dioxide and other factors have interacted to contribute to its increase.
4. Ashe juniper produces flowers during the winter, matures fruits during summer and early fall and disperses seeds in fall and winter. At 1.5 m height (10 to 20 years of age) seeds are produced and as plants increase in size they regularly produce large quantities of seeds.
5. Fruits (seeds) are consumed by nearly all birds and mammals, including domestic livestock, and generally pass unaltered, except for berry removal from the seeds through the digestive tract and can be dispersed in the feces some distance from the source plant. Overland flow of water can also disperse large quantities of seeds.
6. Seed longevity is limited and by the end of two years few seeds remain viable in the field. They are longer lived under laboratory conditions
7. Seedling establishment appears to be possible on nearly all sites and in almost all plant communities. Continuous deep soil with a vigorous, continuous herbaceous plant community can slow the rate of ingress but apparently not prevent it, particularly if periodic fires and/or intensive browsing does not occur. It appears that protection from large herbivores and the presence of high grass cover increases the survivorship of emerged seedlings.
8. Most seedlings occur under mature, large female trees either because of a favorable establishment environment or reduced herbivory impacts.
9. The canopy cover and density of Ashe juniper is significantly influenced by grazing history. Intense browsing by goats, deer and other wildlife can reduce plant numbers and maintain plants in smaller size classes for a given age compared to unbrowsed plants. Communities that are not browsed or grazed frequently have the highest densities and cover of woody plants.
10. Increased size and numbers of Ashe juniper plants in a community greatly alters the production, composition, and diversity of other woody and herbaceous species. Canopy shade and the accumulation of deep litter are primary contributors to these influences. Leaf litter primarily exerts a physical (moisture limiting) affect on other species and not a chemical (allelopathic) effect.
11. Increase in juniper dominance is associated with a threshold where return to grassland dominance becomes difficult and expensive after approximately 25-50 years, depending upon grazing and weather patterns. Managing juniper plants before they produce seeds is an important management strategy.
12. This can be accomplished in many ways (see later chapters) but fire seems the most reasonable ecological and economical method. This, of course, has to be integrated with proper grazing management strategies.
13. Managing Ashe juniper at a landscape level to provide its values where desired and to reduce its impacts where needed is the preferred, though difficult, approach to its management.
Literature Cited
Adams, R.P. and B.L. Turner 1970. Chemosystematic and numerical studies of natural population of Juniperus ashei Buch.. Taxonomy 19:728-751.
Blomquist, K.W. 1990. Selected life history and synecological characteristics of Ashe juniper on the Edwards Plateau of Texas. M.S. Thesis, Dep. Rangeland Ecology and Management, Texas A&M Univ., College Station, TX.
Bray, N.L. 1904. The timber of the Edwards Plateau of Texas: its relation to climate, water supply and soil. U.S. Department of Agriculture, Bureau of Forestry Bulletin No. 49. 30 p.
Brown, D.E. (Editor) 1982. Biotic Communities of the American Southwest – United States and Mexico. Desert Plants 4:1-342.
Bryant V.M. and H.J. Shafer 1977. The late Quaternary paleoenvironment of Texas: A model for the archeologist. Bulletin of Texas Archeological Society 48:1-25.
Cartwright, W.J. 1966. The cedar choppers. Southwestern Historical Quarterly 70:247-255.
Chavez-Ramirez, F. 1992. The role of birds and mammals in the dispersal ecology of Ashe juniper on the Edwards Plateau, Texas. M.S. Thesis, Dep. Wildl. Fish. Sci., Texas A&M Univ, College Station, TX.
Chavez-Ramirez, F. and R.D. Slack 1994. Effects of avian foraging and post-foraging behavior on seed dispersal patterns of Ashe juniper. Oikos 71: 40-46.
Chavez-Ramirez, F. and R.D. Slack 1993. Carnivore fruit-use and seed dispersal of two selected plant species of the Edwards Plateau, Texas. Southwestern Naturalist 38:141-145.
Correll D.S. and M.C. Johnston 1970. Manual of Vascular Plants of Texas. Texas Research Foundation, Renner, TX. 1881 p.
Dalrymple, R.L. 1969. Prescribed grass burning for Ashe juniper control. Progress Report, Noble Foundation, Inc. Ardmore OK. 2p.
Emanuel, W.R., H.H. Shugart, and M. Stevenson 1985. Climatic change and the broad-scale distribution of terrestrial ecosystem complexes. Climate Change 7:29-43.
Foster, J.H. 1917. The spread of timbered areas in central Texas Journal of Forestry 15:442-445.
Fuhlendorf, S.D. 1992. Influence of age/size and grazing history on understory relationships of Ashe juniper. M.S. Thesis, Dep. Rangeland Ecology and Management, Texas A&M Univ., College Station, TX.
Fuhlendorf, S.D. and F.E. Smeins 1997. Browsing and tree size influence on understory of Ashe juniper. Journal of Range Management (in press).
Gehlbach, F.R. 1988. Forests and woodlands of the Northeastern Balcones Escarpment. pp. 27-77 In. Edwards Plateau Vegetation – Plant Ecological Studies in Central Texas. Edited by B.B. Amos and F.R. Gehlbach. Baylor Univ. Press, Waco. 144p.
Hall, M.T., J.F. McCormick and G.G. Fogg 1961. Hybridization between Juniperus ashei, Buckholz and Juniperus pinchotii Sudworth in southwestern Texas. Butler University Botanical Studies 14:9-28.
Holthuijzen, A.M.A. and T.L. Sharik 1984. Seed longevity and mechanisms of regeneration of eastern red cedar (Juniperus virginiana L.). Bulletin of Torr. Botany Club 2:153-158.
Huss, D.L. 1954. Factors influencing plant succession following fire in Ashe juniper woodland types in Real county, Texas. M.S. Thesis, Dep. Rangeland Ecology and Management, Texas A&M Univ., College Station, TX.
Kinucan, R.J. and F.E. Smeins 1992. Soil seed bank of a semi-arid grassland under three long-term (36 years) grazing regimes. American Midland Naturalist 128:11-21.
Kroll, J.C. 1980. Habitat requirements of the Golden-cheeked warbler: Management implications. Journal of Range Management 33:60-65.
Marshall, S. B. 1995. Biogeochemical consequences of livestock grazing in a juniper-oak savanna. M.S. Thesis, Dep. Rangeland Ecology and Management, Texas A&M Univ., College Station, TX.
Marsh, W.M. and Marsh, N.L. 1993. Juniper trees, soil loss, and local runoff processes. (eds.) C.M Woodruff, W.M. Marsh and L.P. Wilding. In Soils, Landforms, Hydrologic Processes, and Land-use Issues-Glen Rose Limestone Terrains, Barton Creek Watershed, Travis County, Texas. Field Report and Guidebook. Soc. Independent Prof. Earth Scientists, Central Texas Chap., Austin, TX.
Merrill, L.B. and V.A. Young 1959. Effect of drought on woody plants. Texas Agriculture Progress 3:9-10.
Owens, M.K. and T.G. Schliesing 1995. Invasive potential of Ashe juniper following mechanical disturbace. Journal of Range Management.
Reinecke, R.K. 1996. Ashe juniper seed production and germination, seedling dynamics and response of live oak/ juniper mottes to summer fire. M.S. Thesis, Dep. Rangeland Ecology and Management, Texas A&M Univ., College Station, TX.
Riskind, D.H. and D.D. Diamond 1988. An Introduction to Environments and Vegetation. pp. 1-15. In. Edwards Plateau Vegetation – Plant Ecological Studies in Central Texas. Edited by B.B. Amos and F.R. Gehlbach. Baylor Univ. Press, Waco. 144p.
Rudloff, E.V. 1968. Gas-liquid chromatography of terpenes. Part XVI. The volatile oil of the leaves of Juniperus ashei Buchholz. Canadian Journal of Chemistry 46:679-683.
Rykiel, E.J. and T.L. Cook 1986. Hardwood red cedar clusters in the postoak savanna of Texas. Southwestern Naturalist 31:73-78.
Smeins, F.E. 1980. Natural role of fire on the Edwards Plateau. pp. 4-16. In Prescribed burning of the Edwards Plateau of Texas. Edited by L.D. White, Texas Agricultural Extension Service, College Station, TX. 74p.
Smeins, F.E. and L.B. Merrill 1988. Long-term Change in a Semi-arid Grassland. In. Edwards Plateau Vegetation – Plant Ecological Studies in Central Texas. Edited by B.B. Amos and F.R. Gehlbach. Baylor Univ. Press, Waco. 144p.
Texas State Soil and Water Conservation Board 1991. A comprehensive study of Texas watersheds and their impacts on watter quality and water quantity. Texas State Soil and Water Conservation Board, Temple TX.
Van Auken, O.W. 1988. Woody Vegetation of the Southeastern Escarpment and Plateau. pp. 43-56 In. Edwards Plateau Vegetation – Plant Ecological Studies in Central Texas. Edited by B.B. Amos and F.R. Gehlbach. Baylor Univ. Press, Waco. 144p.
Van Auken, O.W. 1993. Size distribution patterns and potential population change of some dominatn woody species of the Edwards Plateau region of Texas. Texas Journal of Science 45: 199-210.
Vines, R.A. 1960. Trees, Shrubs and Woody Vines of the Southwest. University of Texas Press, Austin 1104 p.
Weniger, D. 1984. The Explorer’s Texas – The Lands and Waters. Eakin Press, Austin. 224 p.
Wink, R.L. and H.A. Wright 1975. Effects of fire on a juniper community. Journal of Range Management 26:326-329.
Wright, H.A., F.M. Churchill, and W.C. Stevens 1975. Effects of prescribed burning on sediment, water yield, and water quality from dozed juniper lands in Central Texas. Journal of Range Management 29:294-298.
Yager, L. Y. 1993. Canopy, litter and allelopatic effects of Ashe juniper (Juniperus ashei, Buchholz) on understory vegetation. M.S. Thesis, Dep. Rangeland Ecology and Management, Texas A&M Univ., College Station, TX. 109 p.
APPENDIX
| Length | 1 meter (m) = 3.28 feet |
| 1 centimeter (cm) = 0.394 inches | |
| 1 millimeter (mm) = 0.0394 inches | |
| Area | 1 square meter (m2) = 10.76 square foot |
| 1 hectare (ha) = 2.47 acres | |
| Weightper area | 1 kilogram per hectare (kg/ha)= 0.893 pounds per acre |
1. 1 Taxonomic nomenclature follows Hatch et al. (1990)
2. All data are presented in metric units; See the appendix at end of chapter for conversion to english units.
3. Other names applied to the species: Post Cedar, Rock Cedar, Mexican Juniper, Mountain Cedar, Blueberry Juniper (Cedar), Texas Cedar, Sabino, Enebro, Tascate, Taxate, Cedro.
4. Taxonomic nomenclature follows Ransom (1981).
Comments: Allan McGinty, Professor and Extension Wildlife Specialist
Pingback: With Winter Comes Cedar Fever - Nature Fix